
-
摘要: 心肌细胞的微管对维持心肌细胞的正常结构、功能有着密切的关系。而微管的稳定性主要是受微管相关蛋白(MAPs)调节,包括MAP2、MAP4及tau等。翻译后修饰(PTMs)对MAP蛋白的表达和功能的正常发挥有着重要作用。在心力衰竭(心衰)情况下,通常会发生不同程度异常的PTMs,通过调控这些异常的修饰作用可以对心衰进行干预和治疗。因此,本文从心肌细胞微管的功能、微管蛋白的调控等方面出发,对近年来国内外的心衰与微管调控相关研究进行总结,以期为心衰治疗提供理论参考。Abstract: Microtubules in cardiomyocytes have been found to be closely related to the maintenance of normal structure and function of cardiomyocytes. The stability of microtubules is mainly regulated by microtubule-associated proteins(MAPs), including MAP2, MAP4 and tau. Post-translational modifications(PTMs) play an important role in the normal expression and function of MAP proteins. In heart failure, varying degrees of abnormal PTMs usually occur, and it has been shown that intervention and treatment of heart failure can be achieved by modulating these abnormal modifications. Therefore, in this article, we summarize the studies on heart failure and microtubule regulation in recent years from the perspective of the function of microtubules in cardiomyocytes and the regulation of microtubule proteins at home and abroad, in order to provide some theoretical references for the treatment of heart failure.
-
Key words:
- heart failure /
- microtubules /
- tubulin /
- post-translational modifications
-
心力衰竭(心衰)是各种心脏结构或功能异常性疾病引起的一种临床综合征[1-2]。心衰的发病机制非常复杂,基本为心肌收缩和舒张功能障碍[1]。心衰是各种心脏疾病的晚期表现和转归,临床上可分为急性和慢性2种类型,是一种威胁生命的疾病[3-4]。心脏包含多种细胞类型,包括心肌细胞、成纤维细胞、血管平滑肌细胞、内皮细胞和免疫细胞等,其中心肌细胞约占总细胞数量的30%[5-6]。大多数心脏疾病都伴随着心肌细胞的改变,包括代谢、基因表达的异常以及细胞死亡、细胞外基质的变化以及血管生成等[7-9]。从病理生理学角度来讲,心衰的发生和发展与多种机制相关,包括自身免疫和炎症反应的参与、氧化应激反应、RASS系统及交感神经系统激活和神经内分泌因子等体液因子激活。
近年来,随着分子生物学的不断发展推进了研究者们对心衰发病机制的认识。心肌细胞的收缩主要依赖于肌节(sarcomere)的缩短和延展,而该过程还受到了细胞内微管调控蛋白表达量及其转录后修饰的调控[10]。微管的稳定性主要是受微管相关蛋白(microtubule-associated proteins,MAPs)调节,包括MAP2、MAP4及tau等[11]。翻译后修饰(post-translational modifications,PTMs)对MAP蛋白的表达和功能有着重要的作用[12]。在心衰情况下通常会发生不同程度异常的PTMs,通过调控这些异常的修饰作用已被证明可以对心衰进行干预和治疗。
1. 微管及其调控与心脏疾病
1.1 微管与心脏疾病
微管(microtubule)是真核生物细胞骨架重要成分,由α/β微管蛋白二聚体聚合形成的直径约为25 nm的空心管,它们在细胞内的长度可达数十微米,是细胞骨架丝中最硬的部分[13]。微管的主要功能包括参与维持细胞形态、保障细胞分裂期间的染色体移动以及胞内物质的运输等重要生物学过程[14-15]。在心脏中,微管是心肌细胞骨架的主要力量,对维持心肌细胞生理结构,物质传递都发挥着重要作用。已有相关研究证明心肌细胞的微管与外部信号的机械传导以及跨膜蛋白和离子通道的调节有关,对维持心脏正常功能稳定至关重要[16]。如心肌缺血再灌注时会损伤微管,从而引起心肌细胞内微管的聚合与解聚平衡的破坏,导致微管松散、断裂等影响心肌细胞正常生理功能[17]。在心衰中,收缩功能受损的心肌细胞常常被发现伴有微管的增殖、微管运动方式的改变、微管和中间丝网络密度的改变以及肌节蛋白密度的异常变化等[18-21]。
1.2 微管的调控与心脏疾病
微管的稳定性主要是由MAPs来调节,包括MAP2、MAP4及tau等[11]。主要在心肌细胞中表达的MAP4,参与微管网络的病理致密化和基于微管运输的抑制,MAP4的异常表达常常在心脏疾病患者中检测到[22]。而MAPs的表达和功能通常与微管PTMs密切相关。在心衰中通常会发生不同程度的PTMs,这些修饰通过改变MAP与中间丝的相互作用,从而影响微管的稳定[23-25]。MAP4可被微管亲和力调节激酶4(microtubule-affinity regulating kinase 4,MARK4)磷酸化,从而引起蛋白构象的改变以及与微管的结合,进而影响微管的正常动力功能[26-28]。MAP4的去磷酸化已经在压力过载所引起的心脏肥大猫模型上得到证明,同时MAP4的高表达也在心肌病患者的心脏中被证实[18]。
2. 微管的调控与心衰
2.1 微管的去酪氨酸化与心衰
目前研究最充分的PTM是去酪氨酸化,该修饰主要发生在α-微管蛋白C末端。在心脏中,去酪氨酸化的α-微管蛋白是最普遍的PTM微管蛋白,通过这种修饰,可以帮助微管锚定到肌节,并在心脏收缩期间调节微管屈曲。MAPs的去酪氨酸化正在成为科学家们研究调节心肌细胞收缩机制的重要PTM。α-微管蛋白的去酪氨酸化已经被证明可以促进微管的屈曲、承载。与此同时,去酪氨酸化也已经被证实会直接增加心肌细胞的黏弹性刚度并导致心衰患者心肌细胞的收缩功能障碍。Yu等[29]的研究中,MARK4通过促进小鼠MAP4的磷酸化来调节心肌细胞的收缩性,从而有助于微管蛋白羧肽酶血管抑制素2(vasohibin2,VASH2)进入微管,实现对α-微管蛋白的去酪氨酸化,从而实现对心肌细胞功能的调控。同样,Chen等[18]的研究发现,发生心衰时人的心肌细胞微管和“中间纤维结蛋白”含量增加,同时微管去酪氨酸化水平升高。进一步的实验结果表明,微管去酪氨酸化,会增加心肌细胞刚度及黏弹性阻力,从而负向影响心肌细胞的收缩。而使用去酪氨酸化抑制剂处理心肌细胞后,可以显著降低心肌细胞黏弹性阻力,从而改善心肌收缩功能,实现心功能的改善[18]。
2.2 微管的乙酰化与心衰
最常见的乙酰化PTM是指α-微管蛋白N末端K40处的乙酰修饰基团[30],并且,像去酪氨酸一样,它发生在微管聚合物上[31]。另外,K40乙酰化能改变微管的机械性能并保护它们免受机械断裂[32]。近来,在全蛋白质组的研究中又发现了额外的α-微管蛋白和β-微管蛋白乙酰化位点[33]。此外,有报道β-微管蛋白的Lys252上的乙酰化经乙酰转移酶SAN在体外和体内优先发生在非聚合微管蛋白上[34]。K40的乙酰化主要由α-微管蛋白乙酰转移酶1(ATAT1)介导[35, 36],而去乙酰则由组蛋白去乙酰化酶6(HDAC6)[37]或sirtuin 2(SIRT2)[38]催化。
目前,有研究显示微管蛋白乙酰化和心血管疾病存在着一定的联系。高乙酰化α-微管蛋白发生在几种形式的心肌病中[39]。HDAC6过表达导致α-微管蛋白的去乙酰并增加了聚集形成。在蛋白质病变所致心衰的小鼠模型上,HDAC6对微管蛋白去乙酰的抑制作用已经被证实对心脏自噬通量升高时有一定的保护作用[39]。然而,在心脏纤维化和人心房颤动样本中,乙酰化微管蛋白水平降低[40]。去乙酰酶HDAC6降低了ISO诱导的心脏纤维化小鼠中α-微管蛋白的乙酰化水平。HDAC6的抑制逆转了α-微管蛋白乙酰化并改善了心脏成纤维细胞的增殖[40]。此外,近期一项研究证实了心脏成纤维细胞的初级纤毛中含有乙酰化微管蛋白[41]。上述研究结果提示了微管蛋白乙酰化与微管稳定性、灵活性之间存在着动态及分子联系。上述证据提示HDAC6有可能是一个可行的治疗靶点。
2.3 微管的谷氨酰化与心衰
谷氨酰化是一种进化上相对保守且具有重要功能的PTM,主要是在谷氨酸酶的催化作用下,一个或多个谷氨酸残基加到α/β微管蛋白的羧基端的谷氨酸残基上,形成了谷氨酰化[42]。谷氨酰化最早是在脑中的微管蛋白上被发现的,可以调节微管与各种MAP的相互结合,从而影响微管的稳定性。例如Valenstein等[43]的研究结果表明,多聚谷氨酰化可以调节微管剪切蛋白spastin的活性从而影响微管的动态性。早在2015年Kerr等[44]已经在成人心肌细胞中发现了微管蛋白的谷氨酰化,而在Ki等[45]报道的最新研究中发现,在心脏中CEP41介导的纤毛微管蛋白的谷氨酰化,会诱导内皮细胞通过HIF1A-Aurka-VEGF进行迁移,这可能与一些心血管疾病的病理过程有关。Fan等[46]的研究则表明,CEP4介导的微管蛋白谷氨酰化可刺激血管的生成,这与Ⅱ型糖尿病患者的心血管疾病发展有关。
2.4 其他转录后调控的作用
此外常见的PTM还有甘氨酰化、糖基化等等,这些修饰通过调控MAP的表达从而影响微管的功能,从而参与各种代谢和疾病的进展[12]。在心衰中,通过调控相关MAP的表达来降低微管运动机械阻力,可能是改善心肌收缩能力及治疗心衰的重要潜在手段。
3. 以微管为靶点的心衰治疗
目前通过调节微管以及相关蛋白的PTM实现心衰的治疗和干预的主要包括对于秋水仙碱、微管蛋白-酪氨酸连接酶(tubulin-tyrosine ligase,TTL)、微管亲和调节激酶(microtubule affinity-regulating kinase,MARK)等的使用。首先,秋水仙碱作为一个解聚剂,一直被用作治疗疼痛和肿胀等。但其在调节细胞骨架微管中的作用,使得这种古老的药物在心血管疾病包括心衰的治疗中表现了巨大的潜力。早在2007年,Saji等[47]发现秋水仙碱可以显著降低大鼠心肌细胞微管聚合情况,从而明显抑制心肌细胞的凋亡情况,表明微管解聚剂可能是治疗心脏衰竭的潜在治疗策略。其次,微管蛋白作为一个重要的靶标,微管蛋白-酪氨酸连接酶的过表达成为一种高效地减少微管蛋白去酪氨酸化的治疗途径。过量表达微管蛋白-酪氨酸连接酶已被证明可降低心肌细胞的黏弹性,并且可以改善人衰竭心肌细胞的收缩性能以及小鼠梗死心肌细胞的收缩性能[18, 29, 48-49]。此外,微管的稳定性还可以通过激酶来调控结构性的MAP磷酸化,使其与微管解离,实现微管的稳定。例如利用腺苷类药物或心肌细胞特异性缺失的腺苷激酶类药物可以有效防止心肌细胞微管网络的稳定化、微管网络的去酪氨酸化以及病理性心脏肥大和心脏收缩功能障碍[50-51]。在急性心肌梗死的情况下,通过增加MARK4的活性可以过度磷酸化MAP4,从而导致其从微管中解离出来,增加微管与血管抑制素(VASH)的结合,实现去酪氨酸化和心肌细胞收缩功能障碍的调节[29]。
4. 总结
微管作为细胞转运以及细胞骨架的重要组成部分,涉及了各种代谢途径以及细胞功能。在心脏中,心肌细胞的微管网络具有独特的结构和生理特征,对维持心肌细胞的形态、代谢和功能运动等也发挥着重要的作用。如图 1所示,微管蛋白作为维持微管稳定性的重要调节因子,其PTMs可以实现对微管结构的影响从而影响心肌细胞的物理结构和运动。同时,微管网络的致密化、去酪氨酸化等机械地阻碍了心肌细胞的运动,成为引起心脏衰竭的一个重要原因。因此,通过调节微管蛋白的PTMs,实现对微管结构的调控是降低心衰的新型治疗策略。在对微管结构的调控上,传统抗炎药物秋水仙碱、腺苷类药物以及新型的基因治疗调控方式等均表现出了一定的作用,然而这些药物或方法在运用于心衰的治疗前,仍然需要对相关调控机制如MAP如何特异性靶向微管,如何在器官水平和体内影响心脏功能等,进行更加精细全面系统地探究。调控微管蛋白的表达以及微管结构可以成为心脏疾病治疗的一种重要方法。
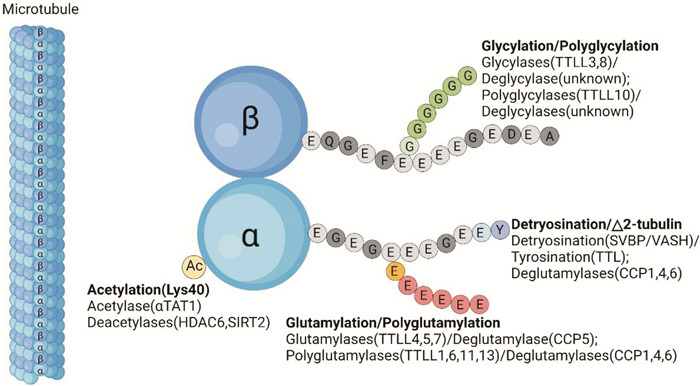 图 1 微管及微管蛋白PTMs的示意图
图 1 微管及微管蛋白PTMs的示意图微管是真核生物细胞骨架重要成分,由α/β微管蛋白二聚体聚合形成的直径约为25 nm的空心管。微管蛋白PTMs发生在微管蛋白的特定部位(乙酰化位于α-微管蛋白的氨基末端结构域)或在C末端尾部内(去酪氨酸化、聚谷氨酰化、聚甘氨酰化),可以调节微管内在性质和MAPs的结合行为。微管的内在动态,加上微管蛋白PTMs和MAP的多尺度调控,赋予了微管群体灵活性和多样性。TTLL:微管蛋白-酪氨酸连接酶样家族;SVBP:血管素结合蛋白;VASHs:血管素;TTL:微管蛋白-酪氨酸连接酶样家族;CCP:胞质羧肽酶样蛋白;αTAT1:α-微管蛋白N-乙酰转移酶1;HDAC:组蛋白去乙酰化酶;SIRT:去乙酰化酶。 Figure 1 Schematic representation of microtubules and PTMs of tubulin利益冲突 所有作者均声明不存在利益冲突
-
[1] McDonagh TA, Metra M, Adamo M, et al. 2021 ESC Guidelines for the diagnosis and treatment of acute and chronic heart failure[J]. Eur Heart J, 2021, 42(36): 3599-3726. doi: 10.1093/eurheartj/ehab368
[2] 程敏, 廖玉华, 袁璟. 2022 ESC速递: 心力衰竭相关临床研究解读[J]. 临床心血管病杂志, 2022, 38(10): 774-776. https://www.cnki.com.cn/Article/CJFDTOTAL-LCXB202210003.htm
[3] Truby LK, Rogers JG. Advanced Heart Failure: Epidemiology, Diagnosis, and Therapeutic Approaches[J]. JACC Heart Fail, 2020, 8(7): 523-536. doi: 10.1016/j.jchf.2020.01.014
[4] Tomasoni D, Adamo M, Lombardi CM, et al. Highlights in heart failure[J]. ESC Heart Fail, 2019, 6(6): 1105-1127. doi: 10.1002/ehf2.12555
[5] Meilhac SM, Buckingham ME. The deployment of cell lineages that form the mammalian heart[J]. Nat Rev Cardiol, 2018, 15(11): 705-724. doi: 10.1038/s41569-018-0086-9
[6] Litviňuková M, Talavera-López C, Maatz H, et al. Cells of the adult human heart[J]. Nature, 2020, 588(7838): 466-472. doi: 10.1038/s41586-020-2797-4
[7] Lopaschuk GD, Karwi QG, Tian R, et al. Cardiac Energy Metabolism in Heart Failure[J]. Circ Res, 2021, 128(10): 1487-1513. doi: 10.1161/CIRCRESAHA.121.318241
[8] Whelan RS, Kaplinskiy V, Kitsis RN. Cell death in the pathogenesis of heart disease: mechanisms and significance[J]. Annu Rev Physiol, 2010, 72: 19-44. doi: 10.1146/annurev.physiol.010908.163111
[9] 柯樊, 廖梦阳, 邱志华, 等. β肾上腺素能受体在梗死后心脏重构中作用的研究进展[J]. 临床心血管病杂志, 2021, 37(4): 298-303. doi: 10.13201/j.issn.1001-1439.2021.04.003
[10] Steele DF, Fedida D. Cytoskeletal roles in cardiac ion channel expression[J]. Biochimica Et Biophysica Acta, 2014, 1838(2): 665-673. doi: 10.1016/j.bbamem.2013.05.001
[11] Dehmelt L, Halpain S. The MAP2/Tau family of microtubule-associated proteins[J]. Genome Biol, 2005, 6(1): 204.
[12] Magiera MM, Singh P, Gadadhar S, et al. Tubulin Posttranslational Modifications and Emerging Links to Human Disease[J]. Cell, 2018, 173(6): 1323-1327. doi: 10.1016/j.cell.2018.05.018
[13] Caporizzo MA, Chen CY, Prosser BL. Cardiac microtubules in health and heart disease[J]. Exp Biol Med(Maywood), 2019, 244(15): 1255-1272. doi: 10.1177/1535370219868960
[14] Goldstein MA, Entman ML. Microtubules in mammalian heart muscle[J]. J Cell Biol, 1979, 80(1): 183-195. doi: 10.1083/jcb.80.1.183
[15] Caporizzo MA, Prosser BL. The microtubule cytoskeleton in cardiac mechanics and heart failure[J]. Nat Rev Cardiol, 2022, 19(6): 364-378. doi: 10.1038/s41569-022-00692-y
[16] Steele DF, Eldstrom J, Fedida D. Mechanisms of cardiac potassium channel trafficking[J]. J Physiol, 2007, 582(Pt 1): 17-26.
[17] Goodson HV, Jonasson EM. Microtubules and Microtubule-Associated Proteins[J]. Cold Spring Harb Perspect Biol, 2018, 10(6): a022608. doi: 10.1101/cshperspect.a022608
[18] Chen CY, Caporizzo MA, Bedi K, et al. Suppression of detyrosinated microtubules improves cardiomyocyte function in human heart failure[J]. Nat Med, 2018, 24(8): 1225-1233. doi: 10.1038/s41591-018-0046-2
[19] Zile MR, Green GR, Schuyler GT, et al. Cardiocyte cytoskeleton in patients with left ventricular pressure overload hypertrophy[J]. J Am Coll Cardiol, 2001, 37(4): 1080-1084. doi: 10.1016/S0735-1097(00)01207-9
[20] Witjas-Paalberends ER, Güçlü A, Germans T, et al. Gene-specific increase in the energetic cost of contraction in hypertrophic cardiomyopathy caused by thick filament mutations[J]. Cardiovasc Res, 2014, 103(2): 248-257. doi: 10.1093/cvr/cvu127
[21] Bollen IAE, van der Meulen M, de Goede K, et al. Cardiomyocyte Hypocontractility and Reduced Myofibril Density in End-Stage Pediatric Cardiomyopathy[J]. Front Physiol, 2017, 8: 1103. doi: 10.3389/fphys.2017.01103
[22] Scholz D, Baicu CF, Tuxworth WJ, et al. Microtubule-dependent distribution of mRNA in adult cardiocytes[J]. Am J Physiol Heart Circ Physiol, 2008, 294(3): H1135-H1144. doi: 10.1152/ajpheart.01275.2007
[23] Robison P, Caporizzo MA, Ahmadzadeh H, et al. Detyrosinated microtubules buckle and bear load in contracting cardiomyocytes[J]. Science, 2016, 352(6284): aaf0659. doi: 10.1126/science.aaf0659
[24] Chinnakkannu P, Samanna V, Cheng G, et al. Site-specific microtubule-associated protein 4 dephosphorylation causes microtubule network densification in pressure overload cardiac hypertrophy[J]. J Biol Chem, 2010, 285(28): 21837-21848. doi: 10.1074/jbc.M110.120709
[25] Cheng G, Takahashi M, Shunmugavel A, et al. Basis for MAP4 dephosphorylation-related microtubule network densification in pressure overload cardiac hypertrophy[J]. J Biol Chem, 2010, 285(49): 38125-38140. doi: 10.1074/jbc.M110.148650
[26] Illenberger S, Drewes G, Trinczek B, et al. Phosphorylation of microtubule-associated proteins MAP2 and MAP4 by the protein kinase p110mark. Phosphorylation sites and regulation of microtubule dynamics[J]. J Biol Chem, 1996, 271(18): 10834-10843. doi: 10.1074/jbc.271.18.10834
[27] Trinczek B, Brajenovic M, Ebneth A, et al. MARK4 is a novel microtubule-associated proteins/microtubule affinity-regulating kinase that binds to the cellular microtubule network and to centrosomes[J]. J Biol Chem, 2004, 279(7): 5915-5923. doi: 10.1074/jbc.M304528200
[28] Drewes G, Ebneth A, Mandelkow EM. MAPs, MARKs and microtubule dynamics[J]. Trends Biochem Sci, 1998, 23(8): 307-311. doi: 10.1016/S0968-0004(98)01245-6
[29] Yu X, Chen X, Amrute-Nayak M, et al. MARK4 controls ischaemic heart failure through microtubule detyrosination[J]. Nature, 2021, 594(7864): 560-565. doi: 10.1038/s41586-021-03573-5
[30] L'Hernault SW, Rosenbaum JL. Chlamydomonas alpha-tubulin is posttranslationally modified by acetylation on the epsilon-amino group of a lysine[J]. Biochemistry, 1985, 24(2): 473-478. doi: 10.1021/bi00323a034
[31] Maruta H, Greer K, Rosenbaum JL. The acetylation of alpha-tubulin and its relationship to the assembly and disassembly of microtubules[J]. J Cell Biol, 1986, 103(2): 571-579. doi: 10.1083/jcb.103.2.571
[32] Xu Z, Schaedel L, Portran D, et al. Microtubules acquire resistance from mechanical breakage through intralumenal acetylation[J]. Science, 2017, 356(6335): 328-332. doi: 10.1126/science.aai8764
[33] Choudhary C, Kumar C, Gnad F, et al. Lysine acetylation targets protein complexes and co-regulates major cellular functions[J]. Science, 2009, 325(5942): 834-840. doi: 10.1126/science.1175371
[34] Chu CW, Hou F, Zhang J, et al. A novel acetylation of β-tubulin by San modulates microtubule polymerization via down-regulating tubulin incorporation[J]. Mol Biol Cell, 2011, 22(4): 448-456. doi: 10.1091/mbc.e10-03-0203
[35] Akella JS, Wloga D, Kim J, et al. MEC-17 is an alpha-tubulin acetyltransferase[J]. Nature, 2010, 467(7312): 218-222. doi: 10.1038/nature09324
[36] Shida T, Cueva JG, Xu Z, et al. The major alpha-tubulin K40 acetyltransferase alphaTAT1 promotes rapid ciliogenesis and efficient mechanosensation[J]. Proc Natl Acad Sci U S A, 2010, 107(50): 21517-21522. doi: 10.1073/pnas.1013728107
[37] Palazzo A, Ackerman B, Gundersen GG. Cell biology: Tubulin acetylation and cell motility[J]. Nature, 2003, 421(6920): 230.
[38] North BJ, Marshall BL, Borra MT, et al. The human Sir2 ortholog, SIRT2, is an NAD+-dependent tubulin deacetylase[J]. Mol Cell, 2003, 11(2): 437-444. doi: 10.1016/S1097-2765(03)00038-8
[39] McLendon PM, Ferguson BS, Osinska H, et al. Tubulin hyperacetylation is adaptive in cardiac proteotoxicity by promoting autophagy[J]. Proc Natl Acad Sci U S A, 2014, 111(48): E5178-E5186.
[40] Tao H, Yang J-J, Shi K-H, et al. Epigenetic factors MeCP2 and HDAC6 control α-tubulin acetylation in cardiac fibroblast proliferation and fibrosis[J]. Inflamm Res, 2016, 65(5): 415-426. doi: 10.1007/s00011-016-0925-2
[41] Villalobos E, Criollo A, Schiattarella GG, et al. Fibroblast Primary Cilia Are Required for Cardiac Fibrosis[J]. Circulation, 2019, 139(20): 2342-2357. doi: 10.1161/CIRCULATIONAHA.117.028752
[42] Eddé B, Rossier J, Le Caer JP, et al. Posttranslational glutamylation of alpha-tubulin[J]. Science, 1990, 247(4938): 83-85. doi: 10.1126/science.1967194
[43] Valenstein ML, Roll-Mecak A. Graded Control of Microtubule Severing by Tubulin Glutamylation[J]. Cell, 2016, 164(5): 911-921. doi: 10.1016/j.cell.2016.01.019
[44] Kerr JP, Robison P, Shi G, et al. Detyrosinated microtubules modulate mechanotransduction in heart and skeletal muscle[J]. Nature Communications, 2015, 6: 8526. doi: 10.1038/ncomms9526
[45] Ki SM, Kim JH, Won SY, et al. CEP41-mediated ciliary tubulin glutamylation drives angiogenesis through AURKA-dependent deciliation[J]. EMBO Rep, 2020, 21(2): e48290.
[46] Fan Z, Peng W, Wang Z, et al. Identification of biomarkers associated with metabolic cardiovascular disease using mRNA-SNP-miRNA regulatory network analysis[J]. BMC Cardiovasc Disord, 2021, 21(1): 351. doi: 10.1186/s12872-021-02166-4
[47] Saji K, Fukumoto Y, Suzuki J, et al. Colchicine, a microtubule depolymerizing agent, inhibits myocardial apoptosis in rats[J]. Tohoku J Exp Med, 2007, 213(2): 139-148. doi: 10.1620/tjem.213.139
[48] Caporizzo MA, Chen CY, Bedi K, et al. Microtubules Increase Diastolic Stiffness in Failing Human Cardiomyocytes and Myocardium[J]. Circulation, 2020, 141(11): 902-915. doi: 10.1161/CIRCULATIONAHA.119.043930
[49] Caporizzo MA, Chen CY, Salomon AK, et al. Microtubules Provide a Viscoelastic Resistance to Myocyte Motion[J]. Biophys J, 2018, 115(9): 1796-1807. doi: 10.1016/j.bpj.2018.09.019
[50] Fassett JT, Xu X, Hu X, et al. Adenosine regulation of microtubule dynamics in cardiac hypertrophy[J]. Am J Physiol Heart Circ Physiol, 2009, 297(2): H523-H532. doi: 10.1152/ajpheart.00462.2009
[51] Fassett J, Xu X, Kwak D, et al. Adenosine kinase attenuates cardiomyocyte microtubule stabilization and protects against pressure overload-induced hypertrophy and LV dysfunction[J]. J Mol Cell Cardiol, 2019, 130: 49-58. doi: 10.1016/j.yjmcc.2019.03.015
期刊类型引用(0)
其他类型引用(1)
-
 下载:
下载:
图(1)
计量
- 文章访问数: 2362
- PDF下载数: 1138
- 施引文献: 1
